2025.10.20
ミトコンドリアゲノム編集法によりナスの細胞質雄性不稔の原因遺伝子の機能証明に成功

【本件のポイント】
- mitoTALEN法及びmitoTALECD法を用い、ナスのミトコンドリア遺伝子のゲノム編集に世界で初めて成功した。
- ターゲットにした細胞質雄性不稔(CMS)遺伝子orf218のDNA配列を改変することで、このミトコンドリア遺伝子のノックアウト系統を作出できた。
- orf218のノックアウト系統では雄性不稔性が消失し、形成された花粉により自殖後代を得ることができた。
- ナスのCMSの原因遺伝子は、ミトコンドリアのorf218であることが証明された。
【研究成果の概要】
高等植物には正常な花粉が形成されない「細胞質雄性不稔(Cytoplasmic Male Sterility; CMS)」と呼ばれる形質が存在します。この形質の原因遺伝子はミトコンドリアゲノムに存在する遺伝子であることが多くの植物で示唆されていますが、ほとんどの場合、直接的な証明はなされていません。一般に、ある形質の原因遺伝子を最終的に特定するには、その遺伝子の変異体を得る、あるいはその遺伝子を形質転換により導入して、表現型が変化するか調べるなどの実験を実施する必要があります。しかし植物のミトコンドリアゲノムの遺伝子を直接改変することは不可能でしたし、ミトコンドリアの形質転換の成功例は皆無でした。したがってCMSの原因遺伝子も、長らく「候補」の状態に留まっていました。しかしながら、2019年に初めて、TALEN法を応用した植物のミトコンドリア遺伝子のゲノム編集が成功し、ミトコンドリアゲノムの改変が可能となりました(Kazama et al., 2019)。
現在、ゲノム編集では、CRISPR-Cas9法を用いることが一般的ですが、ガイドRNAをミトコンドリア内部に輸送できないため、この方法はミトコンドリア遺伝子の編集には適用できません。そこでDNA結合モジュールがタンパク質で構成されるTALEにミトコンドリア移行シグナルを付加したmitoTALEが開発され、FokⅠを介したDNA切断による編集法(mitoTALEN法)と、FokⅠの代わりにシチヂンデアミナーゼを用いたC→T(相補鎖ではG→A)の1塩基置換による編集法(mitoTALECD法)が確立されました。
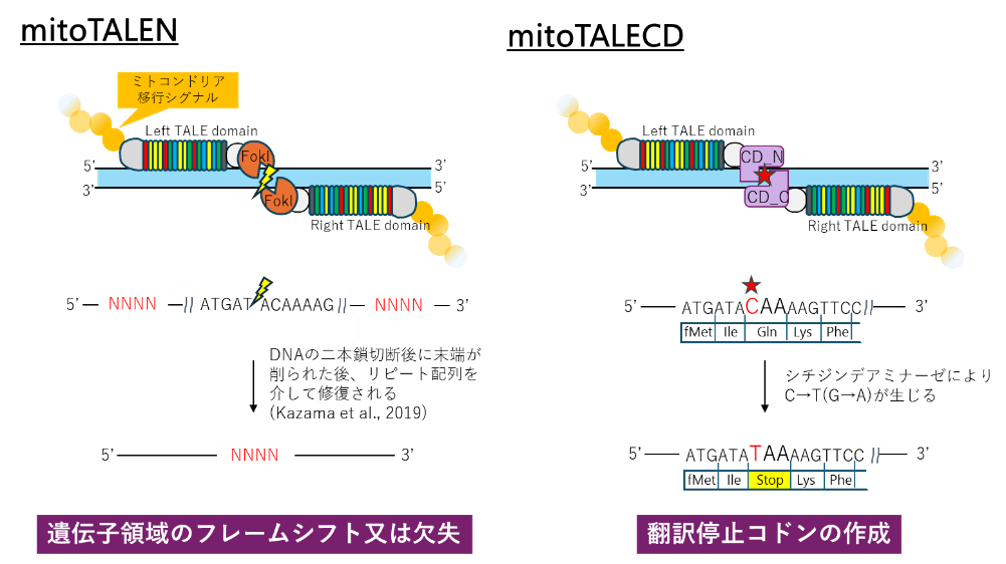
図1 ミトコンドリアゲノム編集法
本研究の材料であるナスには、花粉が形成されないCMS(PN型CMS)が存在します。この原因遺伝子は、いくつかの実験的証拠から、ミトコンドリアゲノムにあるorf218という遺伝子であることが推定されていました(Yoshimi et al., 2013)。今回、このorf218をターゲットに、ミトコンドリアゲノム編集を行いました。
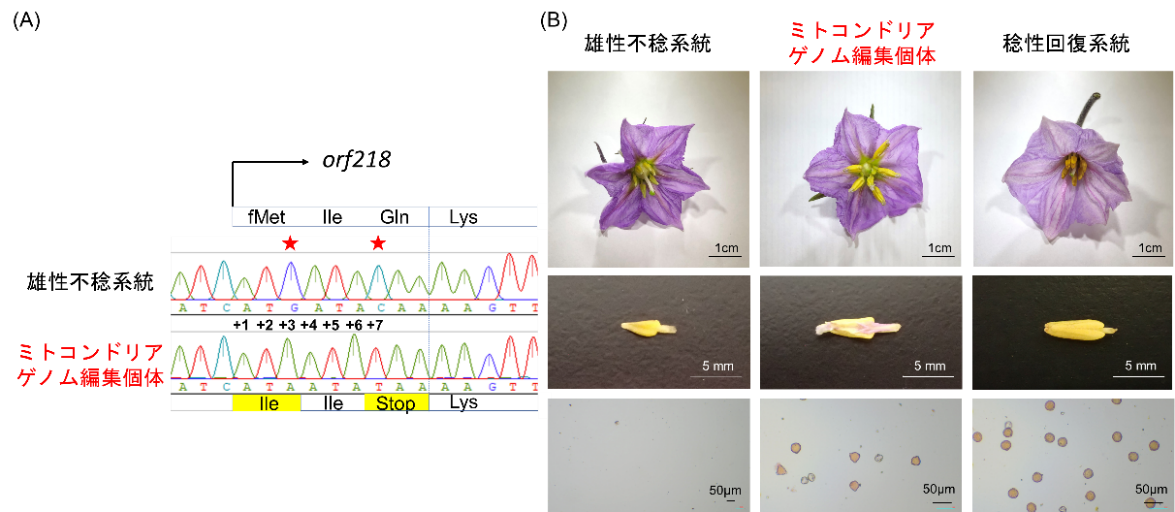
図2 ナスにおけるミトコンドリアゲノム編集
A; ターゲット配列(orf218)内の塩基置換箇所. +3と+7でG→AおよびC→Tの変異が生じている.
B; ミトコンドリアゲノム編集個体の花粉観察結果. 雄性不稔系統では葯内に花粉が存在しないが、編集個体では花粉が認められた.
mitoTALEN法とmitoTALECD法の2つの方法を試した結果、どちらの方法でもミトコンドリアゲノムに改変が生じていることが認められました。特にmitoTALECD法では、得られた編集個体のorf218の塩基配列の10か所に塩基置換が生じていたとともに、開始コドンの消失によってorf218はもはやタンパク質として発現しないことが予測されました。この編集個体を開花するまで育成し、葯の内部を観察したところ、内部に花粉が生じていることが見いだされました。
さらに、この花粉を用いた交配実験では、着果と種子形成が認められました。
以上の結果から、ミトコンドリアのorf218が、ナスにおけるPN型CMSの原因遺伝子であることが証明されました。

図3 ミトコンドリアゲノム編集個体の花粉を使った交配実験
【今後の展開】
今回の実験に用いたPN型CMSでは、雄性不稔性を回復させる稔性回復遺伝子(Rf遺伝子)が別系統のナスの核ゲノムに存在することが明らかになっています。これまでの実験結果から、このRf遺伝子はorf218の転写産物を切断する働きを持っていることが示されています。しかしその実体はまだわかっていません。今回、PN型CMSの原因であると証明されたミトコンドリアorf218と相互作用する核の遺伝子を絞り込むことで、Rf遺伝子の同定が進むことが期待されます。
また、ナスには別種のCMSとして、花粉はできるものの葯が正常に開かない雄性不稔ナス(AI型CMS)が存在します。このAI型CMSについても今回と同様のアプローチで原因遺伝子を特定することができれば、ナスにおけるCMSの全体像を体系的に理解できるものと思われます。
【研究の背景】
CMSは、F1品種の生産に欠かせない形質です。ダイコンやブロッコリーなどのアブラナ科植物や、タマネギ、トウモロコシなど様々な作物でCMSが採種に利用されています。一般的に栽培されているナスもF1品種です。ナスは自殖する植物ですが、一花あたりの種子数が多いため、これまでは人工的な除雄とその後の交配により種子を生産することが可能でした。しかし近年の労働力単価の上昇に伴い、効率的な採種のためにCMSが求められています。また、CMSと、受粉せずに果実肥大が生じる、いわゆる単為結果性とを組み合わせることで、種なしナスの作出も可能となります。このような背景から、我々はナスのCMSの研究を進めてきました。
【研究資金】
本研究は、JSPS科研費20H05680および京都産業大学植物科学研究センター研究費の助成を受けて実施されました。
【論文情報】
論文名:Knocking out of the mitochondrial gene orf218 using mitoTALECD restores
cytoplasmic male sterility in eggplant
和 訳:ナスの細胞質雄性不稔系統では、ミトコンドリアの遺伝子orf218を
mitoTALECD法でノックアウトすると稔性が回復する
著者名:辻村真衣(責任著者)a、静貴子b、宮田暉大c、須佐見朝日c、齊藤 猛雄d、
宮武 宏治d、有村慎一e、山岸博(責任著者)c、寺地徹(責任著者)c
所 属:a龍谷大学農学部、b京都産業大学植物科学研究センター、
c京都産業大学生命科学部、d農研機構、e東京大学農学部
掲載誌:Physiologia Plantarum(John Wiley & Sons社)
公開日:2025年8月9日
U R L:https://onlinelibrary.wiley.com/doi/10.1111/ppl.70446
【問い合わせ先】
<研究に関すること>
龍谷大学農学部 ラボラトリー専門助手 辻村真衣(つじむらまい)(京都産業大学 客員研究員)
Email:mtsukatani@agr.ryukoku.ac.jp
京都産業大学 生命科学部 産業生命科学科 教授 寺地 徹(てらち とおる)
E-mail:terachi@cc.kyoto-su.ac.jp
<取材・報道に関すること>
龍谷大学 農学部教務課
Tel 077‐599‐5601 Email:agr@ad.ryukoku.ac.jp
京都産業大学 広報部
Tel:075-705-1411 E-mail:kouhou-bu@star.kyoto-su.ac.jp